Геномы
В этом разделе описываются генетические характеристики различных живых организмов.
Общие сведения
С парадокс - длина геномов не зависит от сложности организма.
Сравнительные размеры геномов в разных группах организмов.
| Объект | размер генома, пн | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Микоплазмы | 104-106 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Эубактерии (E.coli) | 105-107 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Грибы | (2-5)x107 |
| Вид | Число генов | Длина генома, пн |
| Eubacteria | ||
| Mycoplasma genitalium | 477 | 580.070 |
| Synechocystis sp. | 3168 | 3573 тыс. |
| E.coli | 4280 | 4.639.221 |
| Helicobacter pylori | 1590 | 1667 тыс |
| Bacillus subtilis | 4099 | 4214 тыс |
| Aquifex aelolicus | 1544 | 1551 тыс |
| Micobacterium tuberculosis | 4402 | 4447 тыс |
| Treponema pallidum | 1041 | 1138 тыс |
| Rickettsia prowazekii | 834 | 1111 тыс |
| Thermotoga maritima | 1877 | 1860 тыс |
| Archaea | ||
| Methanococcus sannaschii | 1750 | 1664 тыс |
| Archaeoglobus fulgidus | 2493 | 2178 тыс |
| Aeropyrum pernix | 2620 | 669 тыс |
| Eucaryotes | ||
| Saccharomyces cerevisiae | ~6300 | 12.069 тыс |
| Arabidopsis thaliana | ~26000 | 142.000 тыс |
| Caenorhabditis elegans | ~19000 | 97.000 тыс |
| Drosophila melanogaster | ~14000 | 137.000 тыс |
| X.laevis | ||
| Homo sapiens | ~30000 | 3.200.000 тыс |
Геном эукариот
Геном человека
У человека 23 пары хромосом, 22 аутосомы и 1 пара половых XX (женский пол) или XY (мужской пол).
Гаплоидный геном человека включает более 3 биллионов пар оснований ДНК, общей длинной приблизительно 1,8 м. Полный объем информации записанной в ДНК человека занимает около 750 мегабайт.
Гаплоидный геном человека содержит около 20,000–25,000 генов кодирующих белки.
Около 1.5% генома белок кодирующая, в то время как остальная ДНК включает регуляторные последовательности, интроны, РНК кодирующие последовательности, различные повторы и т.д.
~8% генома приходится на инактивированные последовательности некогда функционировавших ретровирусов HERV (Human endogenous retrovirus), самым молодым из которых, HERV-K, около 5-ти миллионов лет.
Французские исследователи восстановили последовательность одного из ретровирусов семейства HERV-K, содержащиеся в геноме. В клетках линии 239Т происходила транскрипция вирусной ДНК и продукция вирусных частиц. Более того, сам вирус, получивший название Phoenix, оказался способным самостоятельно осуществлять полный ретровирусный цикл от заражения клетки до интеграции в геном и сборки вирусных частиц. В некоторых опухолях, таких как тератокарцинома и меланома, экспрессируются отдельные белки HERV. Этого не достаточно для сборки полноценного вируса - слишком много мутаций. Однако, "воскрешение" полноценного вируса вполне может произойти за счет спонтанной рекомбинации - принципиальная возможность этого подтверждается результатами французских ученых.
| Хромосома | Генов | Длина, пн | Секвенировано |
| 1 | 3,148 | 247,200,000 | 224,999,719 |
| 2 | 902 | 242,750,000 | 237,712,649 |
| 3 | 1,436 | 199,450,000 | 194,704,827 |
| 4 | 453 | 191,260,000 | 187,297,063 |
| 5 | 609 | 180,840,000 | 177,702,766 |
| 6 | 1,585 | 170,900,000 | 167,273,992 |
| 7 | 1,824 | 158,820,000 | 154,952,424 |
| 8 | 781 | 146,270,000 | 142,612,826 |
| 9 | 1,229 | 140,440,000 | 120,312,298 |
| 10 | 1,312 | 135,370,000 | 131,624,737 |
| 11 | 405 | 134,450,000 | 131,130,853 |
| 12 | 1,330 | 132,290,000 | 130,303,534 |
| 13 | 623 | 114,130,000 | 95,559,980 |
| 14 | 886 | 106,360,000 | 88,290,585 |
| 15 | 676 | 100,340,000 | 81,341,915 |
| 16 | 898 | 88,820,000 | 78,884,754 |
| 17 | 1,367 | 78,650,000 | 77,800,220 |
| 18 | 365 | 76,120,000 | 74,656,155 |
| 19 | 1,553 | 63,810,000 | 55,785,651 |
| 20 | 816 | 62,440,000 | 59,505,254 |
| 21 | 446 | 46,940,000 | 34,171,998 |
| 22 | 595 | 49,530,000 | 34,893,953 |
| X | 1,093 | 154,910,000 | 151,058,754 |
| Y | 125 | 57,740,000 | 22,429,293 |
Геном шимпанзе
Геном дрозофилы
Геном нематоды
Геном прокариот
Геном митохондрий
ДНК в митохондриях представлена циклическими молекулами, не образующими связь с гистонами, в этом отношении они напоминают бактериальные хромосомы.
У человека митохондриальная ДНК содержит 16,5 тыс. н.п., она полностью расшифрована. Найдено, что митохондральная ДНК различных объектов очень однородна, отличие их заключается лишь в величине интронов и нетранскрибируемых участков. Все митохондриальные ДНК представлены множественными копиями, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул ДНК. Общее же количество митохондриальной ДНК на клетку составляет около одного процента. Синтез митохондриальных ДНК не связан с синтезом ДНК в ядре. Так же как и у бактерий митохондральная ДНК собрана в отдельную зону – нуклеоид, его размер составляет около 0, 4 мкм в диаметре. В длинных митохондриях может быть от 1 до 10 нуклеоидов. При делении длинной митохондрии от нее отделяется участок, содержащий нуклеоид (сходство с бинарным делением бактерий). Количество ДНК в отдельных нуклеоидах митохондрий может колебаться в 10 раз в зависимости от типа клеток. При слиянии митохондрий может происходить обмен их внутренними компонентами.
рРНК и рибосомы митохондрий резко отличны от таковых в цитоплазме. Если в цитоплазме обнаруживаются 80s рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70s рибосомам (состоят из 30s и 50s субъединиц, содержат 16s и 23s РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50s). В митоплазме на рибосомах идет синтез белков. Он прекращается, в отличие от синтеза на цитоплазматических рибосомах, при действии антибиотика хлорамфеникола, подавляющего синтез белка у бактерий.
На митохондриальном геноме синтезируются и транспортные РНК, всего синтезируется 22 тРНК. Триплетный код митохондриальной синтетической системы отличен от такового, используемого в гиалоплазме. Несмотря на наличие казалось бы всех компонентов, необходимых для синтеза белков, небольшие молекулы митохондриальной ДНК не могут кодировать все митохондриальные белки, только лишь их небольшую часть. Так ДНК размером 15 тыс.н.п. может кодировать белки с суммарным молекулярным весом около 6х105. В это же время суммарный молекулярный вес белков частицы полного дыхательного ансамбля митохондрии достигает величины около 2х106.
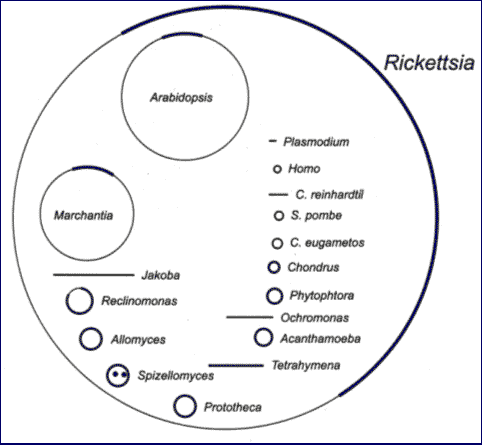
рис. Относительные размеры митохондрий у различных организмов.
Интересны наблюдения за судьбой митохондрий в дрожжевых клетках. В аэробных условиях дрожжевые клетки имеют типичные митохондрии с четко выраженными кристами. При переносе клеток в анаэробные условия (например, при их пересеве или при перемещении в атмосферу азота) типичные митохондрии в их цитоплазме не обнаруживаются, и вместо них видны мелкие мембранные пузырьки. Оказалось, что в анаэробных условиях дрожжевые клетки не содержат полную дыхательную цепь (отсутствуют цитохромы b и a). При аэрации культуры наблюдается быстрая индукция биосинтеза дыхательных ферментов, резкое повышение потребления кислорода, а в цитоплазме появляются нормальные митохондрии.
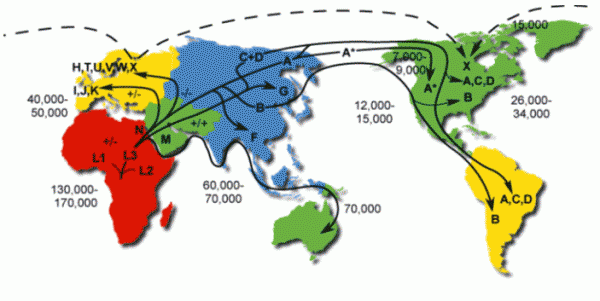
Расселение людей на Земле
Геном пластид
Подобно митохондриям, хлоропласты имеют собственную генетическую систему, обеспечивающую синтез ряда белков внутри самих пластид. В матриксе хлоропластов обнаруживаются ДНК, разные РНК и рибосомы. Оказалось, что ДНК хлоропластов резко отличается от ДНК ядра. Она представлена циклическими молекулами длиной до 40-60 мкм, имеющими молекулярный вес 0,8-1,3х108 дальтон. В одном хлоропласте может быть множество копий ДНК. Так, в индивидуальном хлоропласте кукурузы присутствует 20-40 копий молекул ДНК. Длительность цикла и скорость репликации ядерной и хлоропластной ДНК, как было показано на клетках зеленых водорослей, не совпадают. ДНК хлоропластов не состоит в комплексе с гистонами. Все эти характеристики ДНК хлоропластов лизки к характеристикам ДНК прокариотических клеток. Более того, сходство ДНК хлоропластов и бактерий подкрепляется еще и тем, что основные регуляторные последовательности транскрипции (промоторы, терминаторы) у них одинаковы. На ДНК хлоропластов синтезируются все виды РНК (информационная, трансферная, рибосомная). ДНК хлоропластов кодирует рРНК, входящую в состав рибосом этих пластид, которые относятся к прокариотическому 70S типу (содержат 16S и 23S рРНК). Рибосомы хлоропластов чувствительны к антибиотику хлорамфениколу, подавляющему синтез белка у прокариотических клеток.
 рис.
рис.
Образование шпилек в ДНК некоторых хлоропластов.
Так же как в случае хлоропластов мы вновь сталкиваемся с
существованием особой системы синтеза белка, отличной от
таковой в клетке.
Эти открытия вновь пробудили интерес к теории симбиотического
происхождения хлоропластов. Идея о том, что хлоропласты
возникли за счет объединения клеток-гетеротрофов с прокариотическими
синезелеными водорослями, высказанная на рубеже XIX и XX
вв. (А.С. Фоминцин, К.С.Мережковский) вновь находит свое
подтверждение. В пользу этой теории говорит удивительное
сходство в строении хлоропластов и синезеленых водорослей,
сходство с основными их функциональными особенностями, и
в первую очередь со способностью к фотосинтетическим процессам.

рис. Состав генома пластид у арабидопсиса.
Известны многочисленные факты истинного эндосимбиоза синезеленых
водорослей с клетками низших растений и простейших, где
они функционируют и снабжают клетку-хозяина продуктами фотосинтеза.
Оказалось, что выделенные хлоропласты могут также отбираться
некоторыми клетками и использоваться ими как эндосимбионты.
У многих беспозвоночных (коловратки, моллюски), питающихся
высшими водорослями, которые они переваривают, интактные
хлоропласты оказываются внутри клеток пищеварительных желез.
Так, у некоторых растительноядных моллюсков в клетках найдены
интактные хлоропласты с функционирующими фотосинтетическими
системами, за активностью которых следили по включению С14О2.
Как оказалось, хлоропласты могут быть введены в цитоплазму
клеток культуры фибробластов мыши путем пиноцитоза. Однако
они не подвергались атаке гидролаз. Такие клетки, включившие
зеленые хлоропласты, могли делиться в течение пяти генераций,
а хлоропласты при этом оставались интактными и проводили
фотосинтетические реакции. Были предприняты попытки культивировать
хлоропласты в искусственных средах: хлоропласты могли фотосинтезировать,
в них шел синтез РНК, они оставались интактными 100 ч, у
них даже в течение 24 ч наблюдались деления. Но затем происходило
падение активности хлоропластов, и они погибали.
Эти наблюдения и целый ряд биохимических работ показали,
что те черты автономии, которыми обладают хлоропласты, еще
недостаточны для длительного поддержания их функций и тем
более для их воспроизведения.
В последнее время удалось полностью расшифровать всю последовательность
нуклеотидов в составе циклической молекулы ДНК хлоропластов
высших растений. Эта ДНК может кодировать до 120 генов,
среди них: гены 4 рибосомных РНК, 20 рибосомных белков хлоропластов,
гены некоторых субъединиц РНК-полимеразы хлоропластов, несколько
белков I и II фотосистем, 9 из 12 субъединиц АТФ-синтетазы,
части белков комплексов цепи переноса электронов, одной
из субъединиц рибулозодифосфат-карбоксилазы (ключевой фермент
связывания СО2), 30 молекул тРНК и еще 40 пока неизвестных
белков. Интересно, что сходный набор генов в ДНК хлоропластов
обнаружен у таких далеко отстоящих представителей высших
растений как табак и печеночный мох.
Основная же масса белков хлоропластов контролируется ядерным
геномом. Оказалось, что ряд важнейших белков, ферментов,
а соответственно и метаболические процессы хлоропластов
находятся под генетическим контролем ядра. Так, клеточное
ядро контролирует отдельные этапы синтеза хлорофилла, каротиноидов,
липидов, крахмала. Под ядерным контролем находятся многие
энзимы темновой стадии фотосинтеза и другие ферменты, в
том числе некоторые компоненты цепи транспорта электронов.
Ядерные гены кодируют ДНК-полимеразу и аминоацил-тРНК-синтетазу
хлоропластов. Под контролем ядерных генов находится большая
часть рибосомных белков. Все эти данные заставляют говорить
о хлоропластах, так же как и о митохондриях, как о структурах
с ограниченной автономией.
Транспорт белков из цитоплазмы в пластиды происходит в принципе
сходно с таковым у митохондрий. Здесь также в местах сближения
внешней и внутренней мембран хлоропласта располагаются каналообразующие
интегральные белки, которые узнают сигнальные последовательности
хлоропластных белков, синтезированных в цитоплазме, и транспортируют
их в матрикс-строму. Из стромы импортируемые белки согласно
дополнительным сигнальным последовательностям могут включаться
в мембраны пластиды (тилакоиды, ламеллы стромы, внешняя
и внутренняя мембраны) или локализоваться в строме, входя
в состав рибосом, ферментных комплексов цикла Кальвина и
др.
Удивительное сходство структуры и энергетических процессов
у бактерий и митохондрий, с одной стороны, и у синезеленых
водорослей и хлоропластов – с другой, служит веским аргументом
в пользу теории симбиотического происхождения этих органелл.
Согласно этой теории, возникновение эукариотической клетки
прошло через несколько этапов симбиоза с другими клетками.
На первой стадии клетки типа анаэробных гетеротрофных бактерий
включили в себя аэробные бактерии, превратившиеся в митохондрии.
Параллельно этому в клетке-хозяине прокариотический генофор
формируется в обособленное от цитоплазмы ядро. Так могли
возникнуть гетеротрофные эукариотические клетки. Повторные
эндосимбиотические взаимоотношения между первичными эукариотическими
клетками и синезелеными водорослями привели к появлению
в них структур типа хлоропластов, позволяющих клеткам осуществлять
автосинтетические процессы и не зависеть от наличия органических
субстратов (рис. 236). В процессе становления такой составной
живой системы часть генетической информации митохондрий
и пластид могла изменяться, перенестись в ядро. Так, например
две трети из 60 рибосомных белков хлоропластов кодируется
в ядре и синтезируются в цитоплазме, а потом встраивается
в рибосомы хлоропластов, имеющие все свойства прокариотических
рибосом. Такое перемещение большой части прокариотических
генов в ядро привело к тому, что эти клеточные органеллы,
сохранив часть былой автономии, попали под контроль клеточного
ядра, определяющего в большей степени все главные клеточные
функции.