Клеточная сигнализация
Все клетки способны получать информацию об окружении и изменять свой метаболизм в ответ на поступающие сигналы.
Внеклеточные сигналы могут быть химическими или физическими.
Химические сигналы: аминокислоты и их производные, пептиды, белки, нуклеотиды, жирные кислоты и их производные, стероиды, ретиноиды и малые неорганические молекулы.
Физические сигналы: энергия света, тепло, холод, давление и т.д.
Первичные сигналы воспринимаются рецепторами на клеточной поверхности. Вторичные посредники передают сигнал к клеточным компонентам вовлеченным в ответ. Универсальные вторичные посредники: Ca2+, cAMP (cyclic adenosine monophosphate), cGMP (cyclic guanosine monophosphate), диацилглицерол DG и инозитол трифосфат IP3. Рецепторы воспринимающие сигналы гидрофобных молекул, способных проникать через мембрану
самостоятельно располагаются в цитоплазме.
Свойства сигнальных систем
- Специфичность.
- Адаптация. Есть системы модифицирующие рецепторы (метилирование, рибозилирование). Пример отрицательной обратной связи
- Амплификация. Небольшое число сигнальных молекул способно вызывать реакцию во многих клетках. Один рецептор соединенный с лигандом способен активировать множество G-белков, активирующих вторичные посредники (например, cAMP), которые активируют киназы. Киназы способны активировать множество молекул различных ферментов, запускающих клеточный ответ. Таким образом происходит расширение сигнала
- Интеграция. Усреднение результата
G-белки
G-белки промежуточные компоненты сигнального пути, определяющие будет сигнал усилен или ослаблен. Обычно G-белки состоят из трех субъединиц: a субъединица связывает GDP или GTP. При стимулировании рецептора GDP G-белка заменяется GTP, что приводит
к отделению а-субъединицы, которая связывается и активирует ферменты или ионные каналы. При гидролизе GTP до GDP a-субъединица снова объединяется в комплекс.
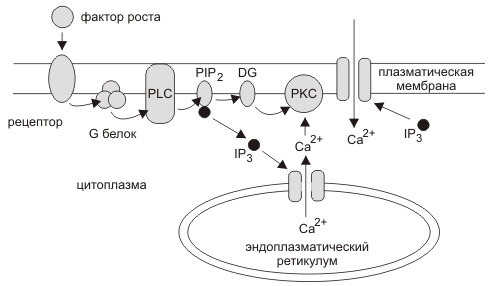
Cигнальный путь вовлекающий вторичные сигналы IP3 и DG. Фактор роста или гормон связывается с рецептором на клеточной поверхности изменяя конформацию рецептора, что стимулирует присоединение GDP к а-субъединице G-белка. а-субъединица диссоциирует от b и g-субъединиц G-белка и GDP меняется на GTP. Активированный G-белок стимулирует фосфолипазу С (PLC), которая гидролизует фосфатидилинозитол 4',5'-бисфосфат (PIP2) до DG и инозитол 1',4'5'-трифосфата (IP3). IP3 активирует ионный канал выпускающий ионы кальция из эндоплазматического ретикулума в цитоплазму. Ионы кальция и DG активируют протеиновые киназы С (PKC) запускающие ответный каскад реакций в клетке.
Другой класс мономерных G-белков состоят из одной субъединицы и называются Ras белками
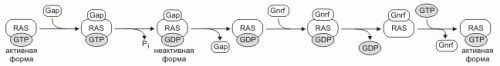
Количество активных Ras определяют GNRF (guanine nucleotide release factors) и GAP (GTPaseactivating proteins). Эти белки обеспечивают обмен GDP на GTP, активируют Ras белки. Гидролиз GTP ингибирует Ras. Некоторые Ras белки негативно регулируются опухолевыми супрессорами.
Киназы и фосфатазы.