Транскрипция у прокариот
РНК-полимераза E.coli
РНК-полимераза E.coli осуществляет транскрипцию всех бактериальных генов и состоит из нескольких субъединиц: α-35кДа, β‘-165кДа, β-155кДа, σ-чаще 70кДа (σ70). РНК-полимераза состава ααββ’σ70 называется holo-фермент (Еσ70), состава ααββ’- core-фермент (E).
σ - сменный фактор специфичности, который диссоциирует после инициации транскрипции. Элонгация и терминация осуществляется core-ферментом. У Е.coli ~10 видов σ-субъединиц. Транскрипция генов теплового шока, оперонов gln или nif осуществляется σ54 в составе holo-фермента Eσ54 (54 кДа).
Все субъединицы заряжены отрицательно: σ>α>β>β’ – расположены по убыванию заряда. В каждой субъединице имеется кластер (+)-заряженных участков, которыми они связываются с ДНК. Наибольшее число кластеров у – β’, который участвует в связывании фермента с ДНК, β-субъединица содержит активные центры - инициации и элонгации, α-субъединицы обеспечивают правильное взаимодействие фермента с промоторами. Рифампицин – блокирует инициацию, стрептолидигин – блокирует элонгацию, что говорит о разнесении активных центров в РНК-полимеразе.
Узнавание и связывание RNA-pol с промотором осуществляется holo-ферментом
Одновременно в клетке присутствует около 7000 молекул РНК-полимеразы. Только holo-фермент обладает высоким сродством к специфической последовательности нуклеотидов - промотору, сродство к остальным случайным последовательностям ДНК у него снижено в 10000 раз. У core-фермента одинаковое сродство к любой последовательности нуклеотидов.
Сам по себе сигма - фактор обладает наименьшим сродством к ДНК по сравнению с другими субьединицами РНК-полимеразы, однако он придает holo-ферменту такую конформацию, которая обладает повышенным сродством к промотору.
Стадии узнавания и связывания, а также инициации осуществляются holo-ферментом. Элонгация и терминация осуществляются core-ферментом.
Две α субъединицы - каркас РНК-полимеразы. К ним крепятся остальные субъединицы.
β' - субъединица отвечает за прочное связывание с ДНК за счет кластера положительно заряженных аминокислот.
В β - субъединице находятся два каталитических центра. Один отвечает за инициацию, а другой - за элонгацию. Один центр работает в holo-, а другой - в core- ферменте.
Инициация транскрипции
РНК-полимераза Ecoli узнает два 6н разделенных 25н
Элонгация транскрипции
Терминация транскрипции
Регуляция транскрипции
Схема негативной индукции Жакоба и Моно
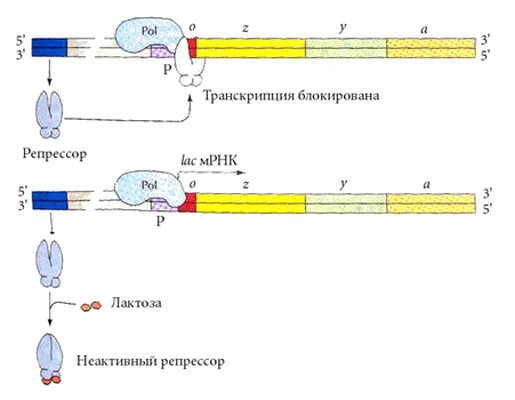
Lac-оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в переносе в клетку дисахарида лактозы и в ее расщеплении.
Z-β - галактозидаза (расщепляет лактозу на глюкозу и галактозу).
Y-β- галактозидпермеаза (переносит лактозу через мембрану клетки).
А - тиогалактозидтрансацетилаза (ацетилирует галактозу).
В отсутствие в клетке лактозы lac-оперон выключен. Активный белок - репрессор, кодируемый в моноцистронном опероне (LacI) , не имеющем оператора, связан с оператором lac-оперона. Поскольку оператор перекрывается с промотором, даже посадка РНК-полимеразы на промотор невозможна.
Как только некоторое количество лактозы попадает в клетку, две молекулы субстрата (лактозы) взаимодействуют с белком - репрессором, изменяют его конформацию - и он теряеет сродство к оператору.
Тут же начинается транскрипция lac-оперона и трансляция образующейся mРНК; три синтезируемых белка участвуют в утилизации лактозы.
Когда вся лактоза переработана, очередная порция репрессора, свободного от лактозы, выключает lac-оперон.
Схема позитивной индукции
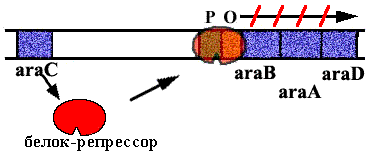
В Аra-опероне E. сoli 3 цистрона, которые кодируют ферменты, расщепляющие сахар арабинозу. В норме оперон закрыт. Белок - репрессор связан с оператором.
Когда в клетку попадает арабиноза, она взаимодействует с белком - репрессором. Белок - репрессор меняет конформацию и превращается из репрессора в активатор, взаимодейсивующий с промотором и облегчающий посадку РНК-полимеразы на промотор.
Эта схема регуляции называется позитивной индукцией, поскольку контролирующий элемент - белок - активатор "включает" работу оперона.
Схема позитивной репрессии
В опероне синтеза рибофлавина у Вacilus subtilis располагаются цистроны ферментов синтеза рибофлавина. Есть белок-активатор, обеспечивающий посадку РНК-полимеразы на промотор. В норме оперон открыт. Образуется N молекул рибофлавина.
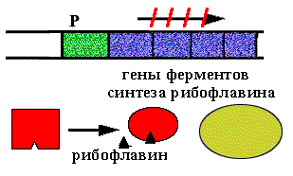
N+1-ая молекула (лишняя) взаимодействует с активатором и он теряет способность активировать посадку РНК-полимеразы на промотор.
Позитивная репрессия, поскольку в регуляции участвует белок - активатор, а сама регуляция заключается в выключении транскрипции.
Схема негативной репрессии
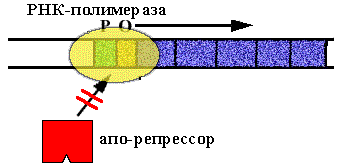
В опероне синтеза триптофана у E. сoli имеется 5 цистронов, которые кодируют ферменты последовательной цепи реакций синтеза триптофана. В норме оперон включен. Белок - репрессор неактивен (в форме апо-репрессора), он не способен садиться на оператор.
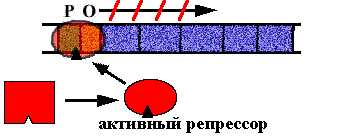
Клетке нужно N молекул триптофана. N+1-ая молекула взаимодействует с апо-репрессором. Он меняет конформацию, садится на оператор и синтез РНК прекращается.
Схема регуляции - негативная репрессия, потому что белок репрессор "выключает" оперон.
Позитивный контроль работы lac-оперона
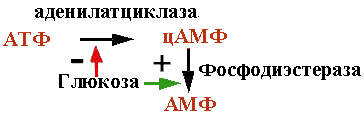
Lac-оперон, подчиняющийся схеме негативной индукции, имеет и позитивный контроль. цАМФ образуется из АТФ ферментом аденилатциклазой. Фосфодиэстераза превращает цАМФ в АМФ. Глюкоза активирует второй и инактивирует первый фермент. Чем больше в клетке глюкозы, тем меньше цАМФ.
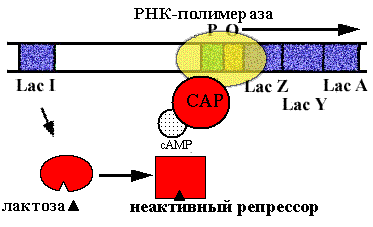
Если нет глюкозы, то цАМФ соединяется с белком катаболической репрессии (САР) и образуется комплекс САР•цАМФ, активирующий посадку РНК-полимеразы на промотор. В присутствии лактозы lac-оперон включается и работает. Если же в клетке есть еще и глюкоза (более экономичный источнок энергии), то нет цАМФ - и активатор не образуется, lac-оперон работает слабо, без дополнительной индукции.